Основы менделевской генетикиЭволюционная экология / Введение / Основы менделевской генетикиСтраница 2
У некоторых организмов встречаются иногда пары аллелей с неполным доминированием. Б этих случаях фенотип гетерозигот 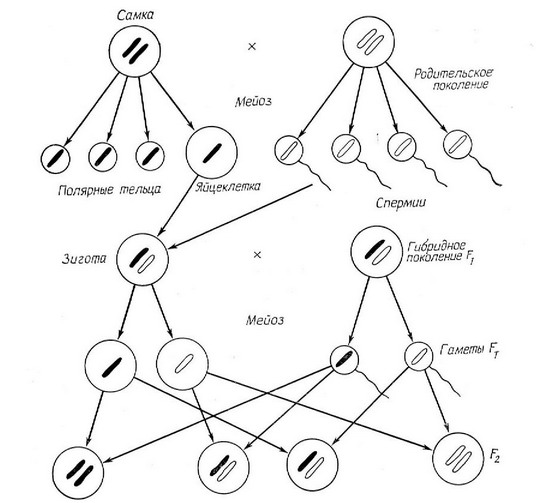
Рис. 1.2. Схема, показывающая, как в процессе деления клеточного ядра геномы родителей распределяются по половым клеткам и снова соединяются посредством рекомбинации во втором (Р) поколении. Для простоты изображена только одна пара хромосом и опущены сложные процессы редукционного деления (мейоза).
занимает промежуточное положение между фенотипами гомозигот, т. е. фенотип в точности соответствует генотипу, и наоборот. По-видимому, аллели, дающие преимущества их обладателям, со временем обычно становятся доминантными, поскольку доминирование обеспечивает использование преимуществ, даваемых данным аллелем, максимальным числом потомков. Очевидная редкость явления неполного доминирования также свидетельствует в пользу того, что в эволюции чаще развивалось полное доминирование. Более того, аллели так называемого дикого типа, преобладающие в природных популяциях, почти всегда доминируют над другими аллелями того же локуса. Генетики предложили многочисленные теории «эволюции доминирования», однако в деталях проблему эту еще нельзя считать разрешенной.
Цитологическое изучение специально приготовленных препаратов клеточных ядер блестяще подтвердило гипотезу Менделя, (рис. 1.2). в ядрах клеток были обнаружены различимые под микроскопом удлиненные плотные образования — хромосомы, содержащие собственно генетический материал — дезоксирибонук-леиновую кислоту (ДНК). У большинства видов ядра диплоидных, клеток, как зиготы (оплодотворенной яйцеклетки), так и соматических (клеток тела), содержат одно и то же число хромосом, хотя у разных видов число это может широко варьировать — от двух у некоторых членистоногих, до сотен у некоторых растений, в диплоидных клетках всегда существуют и обычно четко различаются, пары сходных между собой гомологичных хромосом, однако гаметы содержат лишь половину общего числа хромосом (за исключением гамет полиплоидов), и среди этих хромосом нет гомологичных. Таким образом, гаплоидные клетки содержат только по одному полному набору различных хромосом и аллелей, или по одному геному, тогда как диплоидные клетки содержат двойные наборы. Когда в процессе редукционного деления (мейоза) диплоидные клетки гонад дают начало гаплоидным гаметам, гомологичные хромосомы расходятся по разным гаметам. Нозже, когда гаметы самца и самки сливаются, образуя диплоидную зиготу, дающую в дальнейшем новый диплоидный организм, гомологичные хромосомы снова оказываются объединенными в одной клетке Следовательно, у каждого диплоидного организма один из геномов имеет отцовское происхождение, а другой — материнское Поскольку каждый член пары гомологичных хромосом отделяется от своего гомолога независимо от других пар, хромосомы предыдущего поколения перетасовываются при каждом редукционном делении. Таким образом, генетический материал регулярно перераспределяется и смешивается посредством взаимосвязанных процессов мейоза и слияния гамет.
В каждой хромосоме располагается много различных локусов-и систем аллелей. Два различных признака, контролируемые разными аллелями, локализованными в одной хромосоме, не разделяются строго независимо, а, как показывает статистическая проверка, оказываются связанными друг с другом, демонстрируя явление сцепления генов. Во время мейоза гомологичные хромосомы могут обмениваться своими участками посредством кроссин-говера, в результате чего происходит рекомбинация. Поскольку частота кроссинговера зависит от расстояния между отдельными локусами в хромосоме, генетики могут использовать кроссинговер для «картирования» хромосом. Благодаря тесному сцеплению целые блоки статистически связанных аллелей могут передаваться потомству как функционально интегрированные единицы «коадап-тированных» генов.
Смотрите также
Экономические механизмы охраны природы
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Экологическая ниша
Понятие ниши пронизывает все сферы экологии. Если бы термину «экологическая ниша»
не придавали так много самых разных значений, то экологию можно было бы определить
как науку о нишах. Многие аспек ...
Аутоэкология
Экологию часто разделяют на аутоэкологию и синэкологию. Эти две науки отличаются
предметом и методами. Синэкология является наукой об экосистемах, их структуре и
функционировании. Аутоэкология изу ...