Чистая скорость размножения и репродуктивная ценностьЭволюционная экология / Принципы популяционной экологии / Чистая скорость размножения и репродуктивная ценностьСтраница 1
Очевидно, что немногие организмы реализуют в течение жизни весь свой репродуктивный потеппиал, в связи с чем возникает задача опенить число потомков, произведенных особями, подверженными обычной средней смертности. Для этого используют показатель чистой скорости размножения (К , определяемый как среднее число потомков нулевого возраста, произведенных в среднем организмом за всю его жизнь. Математически Кд — это просто сумма произведений спепифической для каждого возраста выживаемости и плодовитости, где суммирование проводится по всем возрастам, в которых происходит размножение: 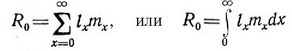
Очевидно, что вместо пределов о и оо могут быть использованы а и ю, так как произведение I равно нулю для любого возраста, в котором не происходит размножения. Подчеркнем еще раз, что левое уравнение соответствует дискретным возрастным классам, а правое — непрерывной градации возрастов. Табл. 5.1 иллюстрирует подсчет Rg по дискретным значениям и т, а на рис. 5.4 графически представлены непрерывно меняющиеся исходные данные.
Популяция растет, если R„>1, стабильна, если R„=l, и сокращается, если Поэтому чистую скорость размножения называют также скоростью замещения популяции. Стабильная равновесная популяция, характеризующаяся круто спадающей L-кри-вой, должна иметь соответственно и высокие значения для того, чтобы заместить саму себя (когда смертность велика, рождаемость также должна быть велика). И наоборот, когда значения высоки, значения от должны быть низки, чтобы поддержать равенство Ro=l. 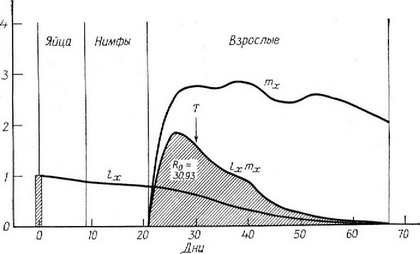
Рис. 5.4. Непрерывное распределение величин и т и их произведений для популяции вши Pediculus humanus в зависимости от возраста. Заштрихованная площадь под кривой соответствует реализованной плодовитости, кото-пая равна чистой скорости размножения (Из Smith F. E. Dynamics of Growth Processes, ed. E. J. Boell (copyright 1954 by Princeton University Press), рис. 1, с. 278, no Evans, Smith, 1952; напечатано с разрешения Princeton University Press.)
Другая важная концепция, разработанная впервые Фишером (Fisher, 1930), касается репродуктивной ценности. Какой вклад в среднем вносят члены определенной возрастной группы в образование тех особей следующего поколения, которые дожили до данного возраста? В стабильной, не увеличивающейся и не уменьшающейся, популяции репродуктивная ценность (у) определяется как специфическое для каждого возраста ожидание будущего потомства. Математически данная величина для стабильной популяции в условиях равновесия определяется следующей формулой: 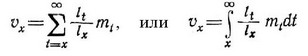
Как и раньше, левое уравнение — для дискретных возрастных групп, а правое—для непрерывного возрастного распределения.
Член представляет собой вероятность выживания особей возраста X до возраста а /и, — средний успех размножения особей возраста 1. Ясно, что для новорожденных особей в стабильной популяции точно равняется чистой скорости размножения Репродуктивная ценность особи, достигшей пострепродуктивного периода, равна нулю, так как от нее нельзя ожидать новых потомков; кроме того, поскольку естественный отбор оперирует только посредством дифференциального успеха размножения (гл. 1), такая закончившая размножение особь не подвержена действию отбора (см. также с. 154—157). При разнообразных значениях 4 и репродуктивная ценность максимальна в начале периода размножения, а затем она уменьшается, так как с возрастом плодовитость обычно падает (хотя сама плодовитость является объектом естественного отбора — см. с. 141 — 154). В табл. 5.1 приведен способ расчета репродуктивной ценности в стабильной популяции. На рис. 5.5 показано, как меняется репродуктивная ценность особей в зависимости от возраста в разных популяциях.
В популяциях с меняющейся численностью репродуктивная ценность представляет собой полученную для настоящего момента оценку потомства, ожидаемого в будущем. Фактически она определяется числом потомков, которых должна была произвести в возрасте х особь, не дожившая до возраста х + 1, для того чтобы оставить столько потомков, сколько она могла бы их произвести, если бы дожила до возраста х+1 и далее вела бы себя как особь, характеризующаяся средней выживаемостью и плодовитостью. В растущей популяции (рис. 5.5, А) репродуктивная ценность очень молодых особей низка по двум причинам. Во-первых, из-за того, что существует некоторая вероятность смерти до наступления репродуктивного периода, и, во-вторых, из-за того, что размножающаяся популяция в будущем станет больше, и относительный вклад будущего потомства в общий генофонд меньше, чем у потомства, родившегося в настоящий момент. 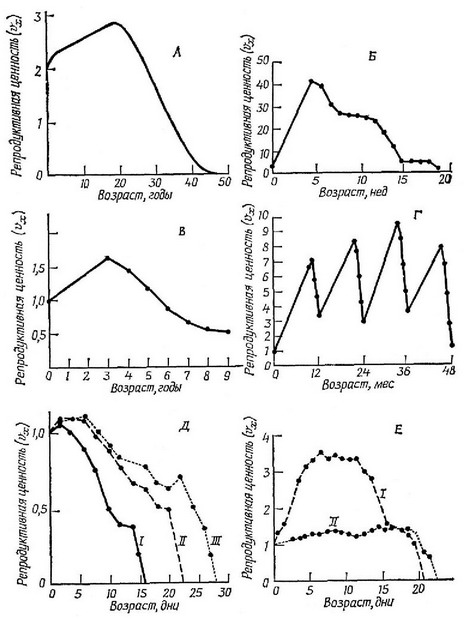
Смотрите также
Аутоэкология
Экологию часто разделяют на аутоэкологию и синэкологию. Эти две науки отличаются
предметом и методами. Синэкология является наукой об экосистемах, их структуре и
функционировании. Аутоэкология изу ...
Заключение.
Основным критерием экологической политики вообще является ее
действенность. Распространено мнение, что в
экологической сфере наиболее действенным является административный подход, то есть подход рас ...
К популяционной организации политипического вида (на примере рыжей полевки -
clethrionomys glareolus shreb.)
Анализ популяционной организации и динамики численности европейской рыжей полевки
- типичного представителя мегаареальных политипических видов млекопитающих Палеарктики
- подтвердил высказанное на ...