Сопряженная эволюция рождаемости и смертностиЭволюционная экология / Принципы популяционной экологии / Сопряженная эволюция рождаемости и смертностиСтраница 1
Очевидно, что вид, характеризующийся высокой смертностью, для того чтобы выжить, должен обладать высокой плодовитостью. Ясно также, что вид, отличающийся повышенной плодовитостью. 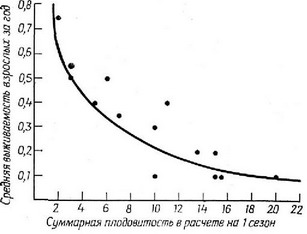
Рис. 5.26. Суммарная плодовитость в расчете на 1 сезон, сопоставленная с выживаемостью в следующем репродуктивном сезоне 14 популяций ящериц. (Из Tinkle, 1969.) Кривая соответствует популяциям, у которых чистая скорость размножения {R) равна единице. Отклонения от этой кривой могут быть связаны как с большей или меньшей величиной Ra, так и с различиями в выживаемости молоди.
должен в среднем претерпевать и значительную смертность, так как в противном случае его популяпия будет продолжать все время расти. При малой плодовитости организмы обычно характеризуются низкой смертностью, так же как высокий уровень выживаемости ведет к снижению плодовитости. На рис. 5.26 показана обратная связь между выживаемостью и плодовитостью для ряда популяпий разных видов ящерип- Рождаемость и смертность эволюпионируют вместе и должны сохранять своего рода равновесие: изменение любой из них неизбежно затрагивает другую.
Удобный показатель способности организма увеличивать свою численность — максимальная мгновенная скорость прироста популяпии {гУ, которая объединяет в себе и рождаемость и смертность. В табл. 5.2 показан диапазон значений, которые может приобретать величина г. Поскольку фактическая мгновенная скорость увеличения популяпии, будучи усредненной за достаточно продолжительный период времени, равна нулю, виды, имеющие высокие значения как, например, Escherichia coll, в меньшей мере реализуют свойственную им максимальную скорость увеличения популяпии, чем виды с низким значением г, как, например. Homo sapiens. Фактически как указывает Смит (Smith, 1954), является мерой «суровости» обычных условий обитания данного организма, и эта величина — один из лучших показателей положения организма в континууме г — .АГ-отбора. Виды, которым свойственны более высокие значения как правило, характеризуются и гораздо большей изменчивостью величины фактической скорости увеличения популяпии г по сравнению с теми видами, которые имеют низкие значения г (см. также рис. 5.14).
Из рис. 5.27 видна обратная отрипательная связь между г и временем генерапии Т. Мелкие организмы обычно имеют значительно более высокие значения чем крупные, что объясняется в первую очередь менее продолжительным временем генерапии, положительная корреляпия которого с размером тела показана на 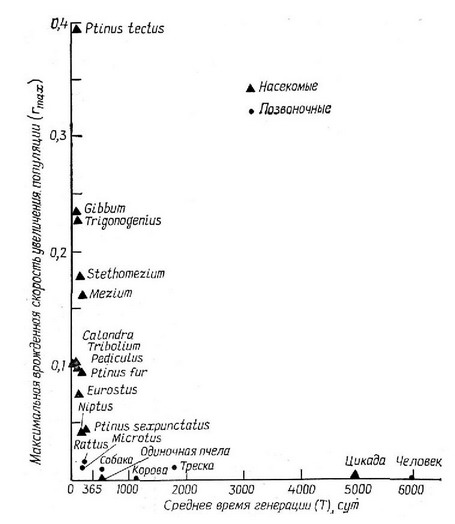
Рис. 5.27. Гиберболическая зависимость между максимальной врожденной ско-роствю увеличения популяции г, и средним временем генерации Т, ввфажен-HBIM в днях. (Из Pianka, 1970; частично заимствовано из Smith, 1954.)
рис. 5.28. Причина подобной корреляции понятна, так как организму, для того чтобы достичь крупных размеров, требуется значительно больше времени. Отсрочка периода размножения также неизбежно ведет к сокрашению
Тем не менее преимушества, даваемые большими размерами тела, должны достаточно часто перевешивать недостатки, связанные с сокрашением так как в противном случае крупные организмы никогда не появились бы в процессе эволюции. Тенденция к возрастанию размеров тела с течением геологического времени, прослеженная по ископаемым остаткам для многих филетических рядов (Newell, 1949), послужила основанием для введения понятия «увеличение филетического размера». Многие преимушества, даваемые крупными размерами тела, совершенно очевидны, но то же самое можно сказать и о связанных с этим 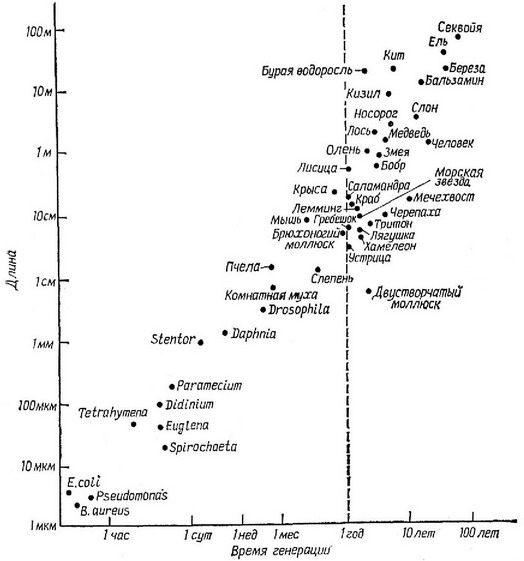
Рис. 5.28. Соотношение длины тела и времени генерации (обе величины в логарифмическом масштабе) в пределах широкого круга организмов. (Из John Tyler Bonner, Size and Cycle: An Essay on the Structure of Biology, Copyright 1965 by Princeton University Press, рис. 1, стр. 17. Перепечатано с разрешения Princeton University Press.)
недостатках. Конечно, более крупный организм имеет меньше
Смотрите также
Платежные инструменты
Платежные инструменты - это правовые средства осуществления переводов денежных средств (наличных и безналичных) с целью совершения платежа.
Порядок выполнения операций с применением платежных инст ...
Структура сообщества
За исключением краткого рассмотрения биомов в гл. 3, до сих пор мы касались только
экологии особей и популяций. В этой главе речь пойдет об экологии сообществ. Подобно
тому как популяции обладают ...
Биогеография морского бентоса
в биосфере Земли можно выделить четыре основных типа сравнительно независимых
друг от друга ЦС: морские, пресноводные и наземные. В свою очередь морские ЦС можно
разделить на бентосные, биотоп кот ...