ВыводыЭкология морского бентоса / Синэкология / ВыводыСтраница 1
Приведённые выше примеры показывают, что для образования очага эндемизма вовсе не обязательны ни физические преграды, ни продолжительное время: он возникает тогда, когда возникают новые экосистемы на новых площадях, которые не могли быть населены другими экосистемами. Причины этой невозможности самые разные: как непосредственная изоляция новой территории (острова), так и возникновение иных условий, далеко выходящих за те пределы, в которых исходные экосистемы могут нормально функционировать (вершины африканских гор, крупные озёра, Паратетис, Арал, канская флора). Во всех показанных случаях на некоей площади появляются неиспользуемые ресурсы (часто неправильно называемые пустыми нишами). Для использования таких ресурсов и образуются эндемичные для данной площади виды (симпатрично или аллопатрично). Это именно тот процесс, который M.L. Rosenzweig (1978, 1995) назвал competitive speciation.
T.F. Stuessy и др. (1990) предложили процесс симпатрического видообразования назвать «cladogenesis» в том случае, когда исходный вид, дав пару (или более) симпатрич-ных видов, вымер, и «anacladogenesis», если исходный вид сохранился вместе с производным(и) от него; преобразование исходного вида в новом для него месте в другой (без изменения числа видов) они назвали «anagenesis». Cladogenesis и anacladogenesis Е.Ф. Гурьянова называла «букетным видообразованием». Доля видов, образовавшихся в результате этих процессов, различна в разных районах, но в общем имеющиеся данные позволяют сформулировать гипотезу о том, что (1) ранг и доля эндемов тем выше, чем больше отличаются условия в ареале новой экосистемы от условий в ареале экосистемы-донора; (2) доля эндемов тем выше, чем большие затруднения виды испытывают в проникновении в ареал формирующейся экосистемы.
В процессе формирования новых экосистем в условиях малого начального таксономического разнообразия многие таксоны получают возможность освоить ресурсы, обычно им недоступные. При этом часто возникают таксоны, имитирующие таксоны, обычно потребляющие эти ресурсы (параллелизм в развитии), примеры которых имеются во всех учебниках по эволюции. Сходство таких таксонов тем больше, чем на более сходной генетической основе они возникают. Строго говоря, сама возможность имитации обусловлена наличием сходных генотипов. Koгдa такой возможности нет, сходные функции в экосистеме выполняют совсем неродственные таксоны. Например, в Новой Зеландии сформировалась разнообразная фауна гигантских нелетающих кузнечиков (weta) — специализированных распространителей семян растений с сочными плодами (эту нишу обычно занимают млекопитающие, отсутствующие в Новой Зеландии). Kyзнeчики охотно поедают различные плоды, и часть семян при этом проходит через пищеварительный тракт неповреждёнными, их всхожесть даже немного улучшается (Duthie et al., 200б). В отдельных случаях для потребления новых для таксона ресурсов возникают весьма отклоняющиеся морфологически формы («запредельная специализация»). Некоторые примеры запредельной специализации приведены выше, дополнительные примеры можно найти у С.С. Шварца (1980). Запредельную специализацию, по-видимому, можно рассматривать как ароморфозы в понимании А.Н. Северцова.
Исследуя неравномерность эволюции в палеонтологической летописи, надо объяснять не только и даже не столько причины быстрой эволюции в отдельные периоды, сколько причину того, что обычно скорость эволюции гораздо меньше той, которая возможна. Поскольку большой разброс в скоростях видообразования характерен для самых различных таксономических групп, его невозможно объяснить неодинаковыми филогенетическими возможностями разных таксонов, причину столь сильно отличающихся скоростей эволюции следует искать вне эволюционирующего объекта, в его окружении. Наиболее правдоподобным кажется объяснение, что скорость эволюции регулирует биотическое окружение, т. е. экосистема. Вид, входящий в экосистему, потребляет в ней некоторые ресурсы (занимает в ней определённую нишу) и, следовательно, находится под действием стабилизирующего отбора. Стабильность свойств вида, в свою очередь, повышает стабильность сообщества, в которое он входит — т. е. процесс имеет положительную обратную связь. Примеры же быстрой эволюции относятся к случаям ослабления или отсутствия такой регуляции, во всех разобранных случаях это происходило при формировании новых экосистем. По-видимому, в таких условиях скорость эволюции приближается к максимально возможной. В.А. Красилов (1969, 1986) предложил назвать биотически регулируемую эволюцию когерентной
(согласованной), а проходящую вне влияния окружающих экосистем — некогерентной.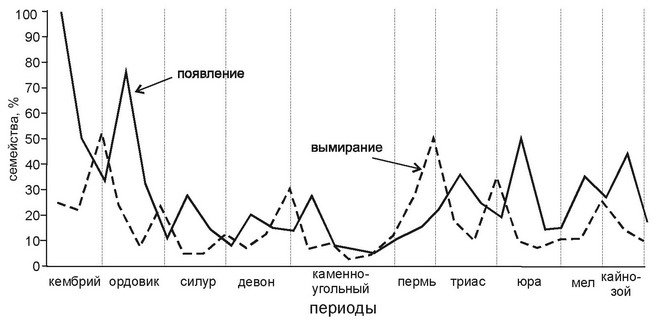
Рис. 4.25.
Вымирание и появление новых семейств у животных. По Newell (1967) из Грант (1980).
Известно, что в истории Земли одни группы сменяют другие: двустворчатые моллюски сменили брахиопод, рептилии сменили земноводных, млекопитаю
Смотрите также
Морфофункциональные особенности лейкоцитов млекопитающих, разводимых в неволе
в условиях европейского севера
Представлены данные о морфофункциональных особенностях лейкоцитов крови различных
видов животных из отряда Carnivora— норок, песцов, лисиц и енотовидных собак, разводимых
в неволе в условиях Карел ...
Дно как биотоп
Люди довольно хорошо представляют себе, как выглядит поверхность суши. Однако
о том, как выглядит поверхность 3/4 Земли — морское дно и какие там условия, у подавляющего
большинства представления ...
Экономические методы охраны окружающей среды и особенности их использования в России
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...