ШельфСтраница 1
Как пишет A. Longhurst (1998), донная фауна Гвинейского залива на поверхности имеет 28°С, а на глубине, не превышающей высоту деревьев тропического леса, всего 16°С. При этом видовой состав биоты края шельфа Гвинейского залива весьма сходен с таковым Западной Европы, а температура отличается не более, чем на 2°С, тогда как видовой состав и абиотические условия верхней сублиторали Гвинейского залива и Западной Европы различаются весьма существенно. А.Н. Голиков указывает, что «северная граница Японской низкобореальной подобласти проходит в районе м. Терпения и северной части о. Итуруп. Однако глубины свыше 30 м в заливах Анива и Терпения занимает Алеутская высокобореальная подобласть» (1982: 97). Тем не менее, несмотря на то, что многочисленные данные свидетельствуют о существовании на шельфе, по крайней мере в умеренных и тропических водах, двух сменяющих друг друга по вертикали биот, районирование проводили и проводят для всего шельфа. Поэтому неудивительно, что рассмотрение всего шельфа как единого биогеографического региона весьма существенно искажает реальную картину. Возможно, пытаясь отразить неоднородность биот по глубине, но оставаясь в рамках «биогеографии всего шельфа) А.Н. Голиков (1980, 1982) вводит в биогеографическую характеристику отдельных видов данные по глубине их обитания, что при выделении действительно био
географических регионов было бы совершенно излишне.
В результате, хотя существуют многочисленные схемы биогеогрфического районирования всего шельфа, нет схем районирования, где бы учитывали биогеографические различия верхней и нижней сублиторали. В первом приближении для обзорных целей можно считать, что фиталь соответствует верхней сублиторали. Поэтому схему районирования по макрофитам можно рассматривать как схему районирования верхней сублиторали. Более того, поскольку в потоках энергии верхней сублиторали существенную роль играет фотосинтез донных макрофитов и рифов герматипных кораллов, эти организмы являются эдификаторами соответствующих ассоциаций, часть из которых — климаксными.
«Литоральная фауна северных берегов Франции и Северного моря идёт на север и восток до Белого моря, мало меняя свой основной состав, но существенно меняется в южной части атлантического побережья Франции. Сублиторальная фауна Северного моря на север доходит только до северных частей Скандинавии, но в нижних слоях мало прогреваемой воды идёт гораздо дальше на юг» (Зенкевич, 1946: 123). 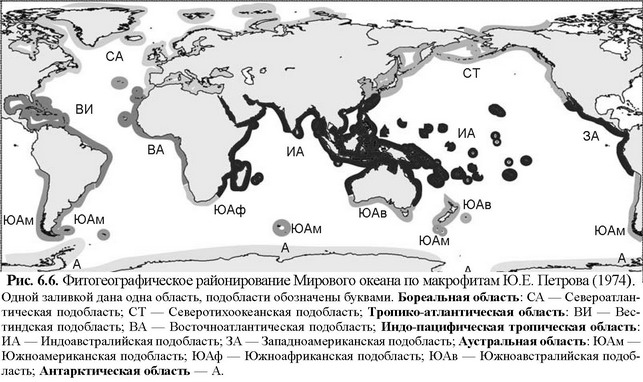
Поэтому схемы районирования по макрофитам и герматипным кораллам должны хорошо соответствовать схеме районирования верхней сублиторали в целом, по крайней мере сообществ твёрдых грунтов.
В первую очередь для карты районирования по макрофитам (рис. 6.6) специфично разделение всей верхней сублиторали на 5 областей: 2 тропических (индо-тихоокеанскую и атлантическую), 2 умеренных (бореальную и аустральную) и антарктическую.
Разделение тропического пояса на два региона подтверждают данные по другой группе-эдификатору верхней сублиторали — герматипным кораллам (см. Главу 3, раздел «Коралловые рифы»).
Как показано в упомянутом разделе, граница между ЦС, эдификаторами которых являются макрофиты и рифы герматипных кораллов, имеет биотическую природу: в более высоких широтах на твёрдых субстратах более конкурентоспособными являются ЦС с климаксом, образованном макрофитами, преимущественно бурыми водорослями, а в низких — герматипными кораллами, на литорали — манграми. При сравнении рис. 6.6 и рис. 3.56 бросается в глаза также совпадение границы тропической зоны с границей распространения рифов герматипных кораллов. Это позволяет сделать вывод, что в верхней сублиторали такое взаимодействие определяет положение границы между тропическими областями и субтропическими регионами. Иными словами, на границе тропиков происходит замена доминирующего таксона климаксных ассоциаций и даже способа фотосинтеза: герматипные кораллы — в тропиках, крупные бурые макрофиты-водоросли — в субтропиках, умеренных и холодных водах. О причинах этого в настоящее время мало что известно. Необходимы исследования как самого симбиоза, так и сравнение биохимии и энергетики фотосинтеза симбиотрофных организмов и макроводорослей, а также биотических взаимоотношений в экосистемах верхней сублиторали. На современном уровне знаний можно отметить следующие моменты:
Смотрите также
Биогеография
Основная цель экологии состоит в том, чтобы понять те разнообразные факторы,
которые влияют на распределение и обилие животных и растений (Andrewartha, Birch,
1954; Krebs, 1972; Mac-Arthur, 1972). ...
Особенности природоохранной деятельности в России.
В нашей стране на первом этапе становления хозяйственного
механизма природопользования недостатки административной системы руководства
проявлялись более явно и отчетливо, чем в других странах. ...
Экономические методы охраны окружающей среды и особенности их использования в России
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...