ВведениеЭкспериментальная генетика и физиология / Популяционные аспекты изучения морфологической изменчивости цестоды proteocephalus
percae (cestoda: proteocephalidea) в онтогенезе / ВведениеСтраница 1
Ранее нами было показано, что эритроциты позвоночных, содержащие гемоглобин с более низким сродством к кислороду (эритроциты барана и быка), имели существенно большую устойчивость к температурному лизису (гемолизу), чем те, в которых гемоглобин находится в высокоаффинной конформации (эритроциты человека, песца и белуги), при том, что красные клетки крови белуги оказались наименее устойчивыми (Борисова, Горюнов, 1997). Для выяснения причин такой взаимосвязи в настоящей работе исследованы параметры реакции автоокисления гемоглобинов и их спектральные особенности и сопоставлены с показателями терморезистентности соответствующих эритроцитов. В основе такого подхода лежит представление о том, что в каскаде реакций автоокисления гемоглобинов образуются свободные радикалы, способные индуцировать процессы повреждения клеточных мембран (Мisга, Fгidоviсh, 1972).
Объектом исследования были гемоглобины человека, песца {Abpex iagopus), барана (Ovis ovis), быка {Bos taurus) и осетра {Acipenser gueldenstaedti). Оксигемоглобин выделяли из свежей крови по стандартной методике; концентрацию белка в растворе определяли на основе коэффициентов экстинкции для оксигемо-глобина £ = 15,37 мМ-1см-1 {в расчете на гем) на длине волны 577 нм {Antonini, Brunori, 1971). Реакцию автоокисления изучали в растворах с концентрацией {1,5-2)х10-6М {в расчете на гем) в аэробных условиях в интервале температур от 20 до 50°С, т.е. ниже температуры денатурации, которая {по результатам собственных измерений методом ДСК) составила для исследованных оксигемоглобинов 66-70,5°С, а для метгемогло-бинов — 59-62°С {Борисова, Горюнов, 1997). Для сопоставления с параметрами термоинду-цированного гемолиза эритроцитов в интервале 50-66°С нам важно было определить скорости автоокисления при максимально возможных температурах из тех, при которых белок еще имеет нативную структуру.
В экспериментах использовались следующие буферные системы: 1) 0,01М натрий-фосфатный буфер + 0,15М МаС!, рН 7,4 {фосфатно-солевой буфер); 2) 0,1М натрий-ацетатный буфер, рН 5,15; 3) 0,1М боратный буфер, рН 9,18; 4) 0,01М натрий-фосфатный буфер, рН 6,2 и рН 6,9. Спектры поглощения регистрировали на спектрофотометре «Sресоrd М 40» {Car! Zeiss, Jena).
В растворе в ходе окисления могут содержаться различные формы гемоглобина {окси-, деокси- и метформы). Концентрация каждого из трех компонентов может быть выражена как функция измеренных поглощений путем решения системы уравнений:
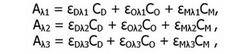
где А — поглощение; Ср, Со и См — молекулярные концентрации деокси-, окси- и метгемогло-бина, соответственно; Л1, Л2, Лз — выбранные длины волн; £Л — коэффициент экстинкции при данной длине волны.
Значения коэффициентов экстинкции для оксигемоглобинов человека были выбраны для длин волн 560, 576 и 630 нм (Веnеsh et а1., 1973; Уап Аssеndеlft, 1970), и после решения системы уравнений определяли количество оксигемоглобинов в реакционной смеси.
Способность разных гемоглобинов к автоокислению характеризовали константой скорости реакции автоокисления:
где М0 и N - концентрации гемоглобина в начальный момент времени t0 и момент времени t, соответственно; К - константа скорости реакции автоокисления.
В ходе эксперимента определяли концентрацию участников реакции автоокисления через определенные промежутки времени и по кинетическим кривым:
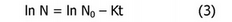
из начальной скорости реакции автоокисления рассчитывали кажущиеся константы скорости реакции.
Поскольку видимые спектры метгемоглобина рН-чувствительны, то для надежного сопоставления способности различных гемоглобинов к автоокислению изучали рН-зависимости скоростей реакции. Для гемоглобинов позвоночных, кроме человека, данные по изменению коэффициентов экстинкции £Л в зависимости от рН отсутствуют, поэтому для определения доли ок-сиформы в реакционной смеси мы пользовались следующей формулой:
Смотрите также
Экологическая ниша
Понятие ниши пронизывает все сферы экологии. Если бы термину «экологическая ниша»
не придавали так много самых разных значений, то экологию можно было бы определить
как науку о нишах. Многие аспек ...
Экономические механизмы охраны природы
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Структура сообщества
За исключением краткого рассмотрения биомов в гл. 3, до сих пор мы касались только
экологии особей и популяций. В этой главе речь пойдет об экологии сообществ. Подобно
тому как популяции обладают ...