Эволюция смертности и старенияЭволюционная экология / Принципы популяционной экологии / Эволюция смертности и старенияСтраница 1
Почему организмы по мере увеличения их возраста стареют? Ведь можно было бы ожидать и прямо противоположного эффекта, поскольку с возрастом организмы становятся опытнее и, следовательно, должны научиться избегать хищников, поскольку у них содержится больше антител и поскольку они вообще становятся умнее и лучше адаптированы как в поведенческом, так и в иммунологическом отношении. Если физиология старения давно интересовала исследователей и ей уделялось должное внимание, то эволюционный аспект данного процесса стали изучать только в самое последнее время (Medawar, 1957; Williams, 1957; Hamilton, 1966; Emlen, 1970). Эта проблема также была предвидена Фишером (Fisher, 1930), который своей концепцией репродуктивной ценности наметил пути ее разработки.
Медавар (Medawar, 1957) хорошо проиллюстрировал эволюцию старения при помощи простой модели, в которой фигурируют неживые объекты. Предположим, что в химической лаборатории
находится запас стеклянных пробирок в количестве 1000 штук, а ежемесячная скорость их разбивания равна 10%. Каждый месяц по совершенно случайным причинам разбивается 100 пробирок, а для того чтобы восполнить их запас, добавляют 100 новых пробирок. (Хотя в данном случае скорость разбивания пробирок — вполне определенная величина, модель может быть легко построена с использованием вероятностных или стохастических понятий.) Все новые пробирки получают отметку о дате начала использования и их возраст (в месяцах) в дальнейшем всегда может быть определен. Каждая пробирка имеет абсолютно одну и ту же вероятность переживания от одного месяца до следующего, равную 900/1000 = 0,9. Таким образом, пробирки, которые изначально были старше, имеют такую же «смертность», как и более молодые, и никакого «старения» не наблюдается: все пробирки потенциально бессмертны. Вероятность выживания в течение двух последующих месяцев представляет собой произведение вероятностей выживания в каждом месяце по отдельности, т. е, 0,9 • 0,9 = 0,9 = 0,81. Вероятность выживания в течение трех месяцев равна 0,9, а в течение X месяцев — 0,9. Спустя несколько лет «популяция» из 1000 пробирок достигнет устойчивого возрастного распределения, при котором 100 пробирок имеют возраст О мес, 90 — 1 мес, 81 — 2 мес, 73 — 3 мес, 28 — 12 мес, ., 8 — 24 мес, ., 2,25 — 36 мес, . не более одной пробирки в данной партии имеет возраст более 48 мес и т. д. (Конечно, данные числа представляют собой просто ожидаемые количества пробирок определенного возраста. Ошибка выборки и стохастические вариации приводят к тому, что числен- 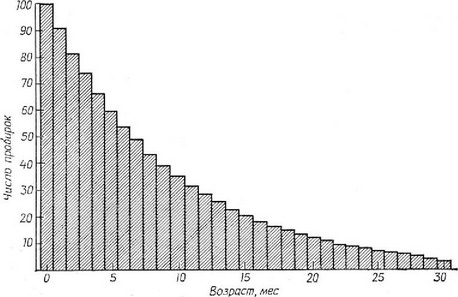
Рис. 5.25. Стабильное возрастное распределение пробирок при скорости их разбивания 10% в месяц. Поскольку очень немногие пробирки «живут» дольше 30 мес, распределение произвольно оборвано на этом возрасте.
ность некоторых возрастных групп будет выше, а некоторых — ниже ожидаемой.) На рис. 5.25 показана часть ожидаемого стабильного возрастного распределения, при котором число более молодых пробирок всегда будет превышать число более старых. Хотя потенпиально каждая пробирка бессмертна, в рассматриваемой группе практически отсутствуют пробирки, возраст которых превышал бы 5 лет. Удлинение срока эксплуатапии пробирки, происходяшее с возрастом, почти неизбежно приводит к ее разбиванию.
Усложняя далее свою модель, Медавар приписал каждой из 900 пробирок, выживаюших в течение 1 мес, одинаковое участие в ежемесячном «размножении» (т. е. в появлении 100 добавляемых вновь пробирок). Таким образом, каждая выживающая пробирка размножается со скоростью 1/9 пробирки в месяп. В этой модели плодовитость не меняется с возрастом, но меняется соотношение размножающихся пробирок. Более молодые возрастные группы участвуют в ежемесячном размножении в значительно большей степени, чем старые, просто потому, что они многочисленнее. Кроме того, спепифическое для их возрастной группы ожидание будущего пополнения или их суммарная репродуктивная пенность также более высоки (рис. 5.25), поскольку у них больше суммарная ожидаемая продолжительность дальнейшей жизни. Однако в такой популяпии, характеризующейся потенпиальной бессмертностью и одинаковой плодовитостью, репродуктивная пенность отдельных индивидуумов с возрастом не меняется.
Теперь, предположив, что наши пробирки имеют «гены», рассмотрим судьбу мутанта, фенотипическое выражение которого проявляется в несколько большей хрупкости по сравнению со средней. Очевидно, что такой ген оказывается вредным для своего носителя, поскольку он снижает вероятность его выживания, а следовательно, и приспособленность. Подобный мутант находится в неблагоприятных с селективной точки зрения условиях, и он неизбежно будет элиминирован из популяпии. На следующем этапе рассмотрим судьбу другой группы мутантных аллелей по локусу, контролирующему время проявления гена хрупкости. Различные аллели этого второго локуса по-разному изменяют время проявления гена хрупкости: некоторые приводят к его сдвигу на более ранний возраст, а другие — на более поздний. Очевидно, что пробирки, несущие ген хрупкости и ген-модификатор, тормозящий его проявление, имеют преимущество по сравнению с пробирками, гены-модификаторы которых приводят к более раннему проявлению гена хрупкости, поскольку они живут в среднем дольше и, следовательно, производят большее число потомков. Таким образом, хотя ген хрупкости постепенно и элиминируется отбором, «поздние» модификаторы накапливаются за счет гибели «ранних» модификаторов. Чем позднее проявляется ген хрупкости, тем больше вклад данного индивидуума в будущие поколения. В предельном случае, когда репродуктивная ценность снижается до нуля, естественный отбор, оперирующий только путем дифференциального успеха размножения, не может далее откладывать проявления губительного признака, эффект которого выражается в старении. Таким образом, признаки, проявление которых посредством отбора генов-модификаторов переносится на период старости, успешно изымаются из генофонда популяции. По этой причине период старости можно рассматривать как своего рода «генетическую свалку», а процесс отбора, откладывающий проявление губительных генетических признаков, называют удалением открытых эффектов аллеля.
Смотрите также
Экономические механизмы охраны природы
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Биогеография морского бентоса
в биосфере Земли можно выделить четыре основных типа сравнительно независимых
друг от друга ЦС: морские, пресноводные и наземные. В свою очередь морские ЦС можно
разделить на бентосные, биотоп кот ...
Биологические опасности, связанные с пищей
Научно-технический прогресс сильно повлиял на сферу производства продуктов питания. Технологическая обработка продуктов, консервирование, рафинирование, длительное и неправильное хранение резко снизил ...