Теория: колебания хищник — жертваЭволюционная экология / Взаимодействия между популяциями / Теория: колебания хищник — жертваСтраница 2
Рис. 6.18. Три типа функциональной реакции. (По Holling, 1959а.)
Рис. 6.19. А. Зависимость суточного потребления коконов насекомого (жертва) одним хищником от плотности коконов для трех мелких млекопитающих, поедающих коконы насекомого (так называемая функциональная реакция). Б. Плотность каждого из этих млекопитающих в зависимости от плотности жертвы (численная реакция хищников). В. Объединенные функциональная и численная реакции видов-хищников, отражающие общую иптепсивпоств ввгеда-ния популяции жертвы, в зависимости от плотности жертвы. (Из Holling, 1959а.) I - Blari-па; II—Peromyscus; III—So-rex\ IV— общие потери. 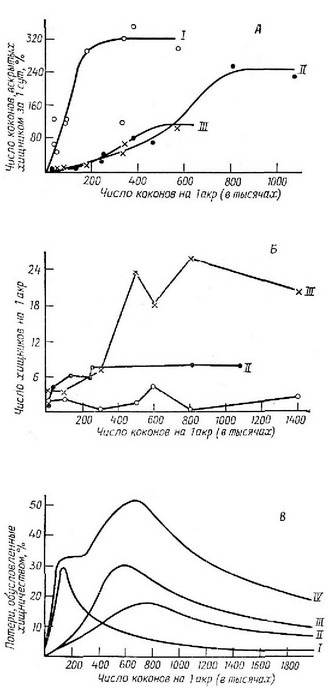 Свежая информация визитки за час у нас.
Свежая информация визитки за час у нас.
менее постоянным (рис. 6.18 и 6.19, А). Во-вторых, возрастание плотности жертвы приводит к увеличению размера популяпии хищника, при этом большее число хищников питается большим числом особей жертвы (рис. 6.19, Б). Первый случай Соломон назвал функпиональной реакцией, а второй — численной реакцией хищника. Различают три отдельных типа функциональной реакции, между которыми возможны переходы (рис. 6.18). [Уравнения (12) и (14) моделируют тип I — линейную функциональную реакцию без верхнего предела.] Обратите внимание, что функциональная реакция хищника допускает регуляцию плотности жертвы без увеличения численности хищника (в отсутствие численной реакции). Используя «системный» подход (см. также гл. 8), который основан на непрерывной обратной связи между наблюдением и моделью, Холлинг (Rolling, 1959а, 1959b, 1966) разработал совершенные модели хищничества, учитывающие как функциональную, так и численную реакцию, а также другие параметры, включая различные временные запаздывания и уровень голодания. Эти модели более реалистичны и точнее описывают события, чем другие (упомянутые выше), но они более сложны и их приложение ограниченно. Очевидно, реалистичная модель взаимосвязи хищник — жертва должна быть довольно сложной!
Простая графическая модель взаимодействия хищник — жертва была разработана Розенцвейгом и Мак-Артуром (Rosenzweig, MacArthur, 1963), которые рассуждали примерно так. В отсутствие хищников максимальная равновесная плотность популяции жертвы равна К„ т. е. предельной плотпости насыщения жертвы. Должен также существовать и нижний предел плотности жертвы, ниже которого контакты между особями становятся слишком редкими, чтобы обеспечить размножение, и численность популяции жертвы падает до пуля. Точно так же и любой данной плотпости жертвы должна соответствовать определенная максимальная плотность хищника, которая может поддерживаться без увеличения или уменьшения популяции жертвы. Исходя из этого, на плоскости Nj—N можно начертить изоклину жертвы (dN,/dt = 0), аналогичную вычерченной на рис. 6.3 и 6.4 (рис. 6.20). Поскольку изоклина жертвы имеет одну-единственную вершину, для выводов. 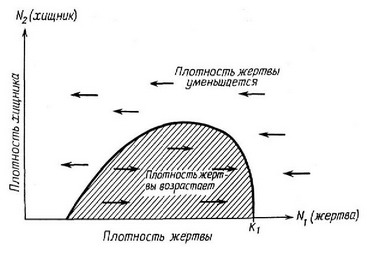
Рис. 6.20. Гипотетическая изоклина вида-жертвы (dN/dt=0), вв1черченпая в координатах плотностей хищника и жертвв:. Популяция жертвв: увеличивается внутри заштрихованной области и уменвшается за ее пределами. При средних величинах нлотности популяция жертвв: характеризуется ввюокой скороствю оборота, что без умепвшепия числеппости ее популяции позволяет хищнику достичв вв1-сокой нлотности. 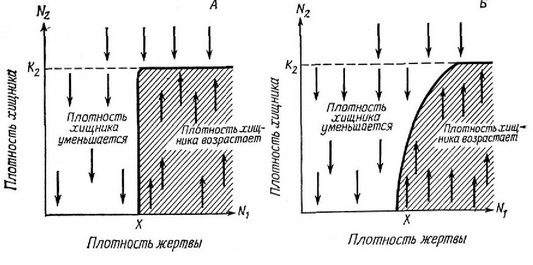
Рис. 6.21. Две гипотетические изоклины хищника. А. Ниже определенной пороговой плотности жертвы X отдельные хищники не могут добывать за единипу времени столько нищи, сколько им требуется для воспроизведения, в области, расположенной на графике левее этой пороговой плотности жертвы, популяпия хищника уменьшается, правее — увеличивается (заштриховаппый участок); это увеличение возможно лишь тогда, когда плотность хищника ниже его предельной плотности насыщения K.j. До тех пор пока хищники не мешают друг другу эффективно добывать нищу, их изоклина идет строго вертикально вплоть до предельной плотности насыщения, как показано на А. Б. Если при высокой плотности хищников конкуренпия между ними снижает эффективность добывания пищи, то их изоклина пойдет с пекоторым наклоном, как показано на рисунке. Быстрое обучение жертвы тактике избегания в результате частых столкновений с хищником будет иметь такие же последствия.
вытекающих из модели, точная форма кривой не имеет значения. Выше этой кривой популяция жертвы уменьшается, ниже — увеличивается. Теперь рассмотрим форму изоклины хищника (dN2/dt = Q) .{.шжс: определенного порога плотности жертвы отдельные хищники не обеспечиваются достаточным количеством пищи, чтобы воспроизвести себе замену, и поэтому популяция хищника должна уменьшаться; выше этого порога количество хищников будет возрастать. Для простоты сначала допустим (ниже это допущение опущено), что взаимодействие или конкуренция между хищниками ослаблены; это сл5ается тогда, когда хищники лимитируются не количеством жертвы, а другими факторами. При этом условии изоклина хищника должна выглядеть так, как она изображена на рис. 6.21, А. Если между хищниками существует конкуренция, то для поддержания более высокой плотности хищника потребуется больше жертв и изоклина будет иметь некоторый наклон, как показано на рис. 6.21, Б. В обоих примерах принято, что предельная плотность насыщения хищника зависит не от плотности жертвы, а от других причин. На плоскости N\ — N устойчивое равновесие обоих видов возможно только в одной точке — точке пересечения двух изоклин (где dN/dt и dN/dt одновременно равны нулю). Теперь рассмотрим поведение этих двух популяций в каждом из четырех квадрантов, обозначенных буквами А, В, Си D на рис. 6.22. В квадранте А плотность обоих видов увеличивается, в В увеличивается плотность хищника, а плотность жертвы уменьшается, в С плотность обоих видов уменьшается, а в D плотность жертвы увеличивается, а плотность хищника уменьшается. ГГа рис. 6.22 стрелками, или векторами, отмечены указанные изменения плотности популяций.
Смотрите также
Органические контаминанты
Летучие органические соединения
Летучие органические соединения (ЛОС) — водные примеси, которые представляют опасность, когда их концентрация достигает даже незначительных уровней. Отличительная особ ...
Структура сообщества
За исключением краткого рассмотрения биомов в гл. 3, до сих пор мы касались только
экологии особей и популяций. В этой главе речь пойдет об экологии сообществ. Подобно
тому как популяции обладают ...
Платежные инструменты
Платежные инструменты - это правовые средства осуществления переводов денежных средств (наличных и безналичных) с целью совершения платежа.
Порядок выполнения операций с применением платежных инст ...