Пирамиды энергии, численностей и биомассыЭволюционная экология / Структура сообщества / Пирамиды энергии, численностей и биомассыСтраница 1
Чем выше трофический уровень, тем меньше скорость потока энергии через него. Объясняется это несколькими причинами. Поскольку эффективность переноса энергии никогда не бывает равной 100%, не вся энергия, содержащаяся в любом данном пищевом объекте, доступна хищнику. Часть ее теряется во время превращения тканей жертвы в ткани хищника, а часть вообще не ассимилируется и проходит через кишечный тракт хищника в неизменном виде, а затем разлагается редуцентами. Эффективность переноса вещества и энергии от жертвы к хищнику нередко бывает ослаблена выработанной жертвой тактикой избегания хищника, например химической защитой растений. Помимо этого, каждая организменная единица (и трофический уровень) тратит часть поступающей энергии на собственные нужды, еще более уменьшая количество энергии, доступной для более высоких трофических уровней. Наконец, при равновесии вся энергия, усвоенная первичными продуцентами, должна быть истрачена и рассеяна в пространстве в виде тепла, т. е. количество энергии, поступившей в систему, должно быть уравновешено количеством, покинувшим систему.
Падение скорости потока энергии при переходе с одного трофического уровня на другой определяет многие свойства сообщества, в том числе общее количество этих уровней и соотношение хищников и жертв. После приведения данных к общим единицам площади и времени экологи подсчитали, что на любой данный трофический уровень поступает примерно 10—20% энергии предыдущего уровня. Следовательно, из 1000 кал, получаемых первичными продуцентами, вторичным хищникам, отделенным тремя трофическими уровнями, достанется лишь небольшая часть. Последствием такого быстрого уменьшения количества энергии является тот факт, что численность животных высоких трофических уровней обычно гораздо меньше, чем животных низких уровней. Кроме того уменьшение количества энергии ставит верхний предел возможному числу трофических уровней, которое обычно равно 5—6.
Удобным способом выражения энергетической структуры сооо-щества является пирамида энергии, которая строится на основе данных по скоростям потока энергии между разными трофическими уровнями (рис. 8.5). Из законов термодинамики и упомянутых выше соображений следует, что. пирамида энергии никогда не может быть перевернутой, т. е. с увеличением номера трофического уровня поток энергии через уровень всегда должен уменьшаться. 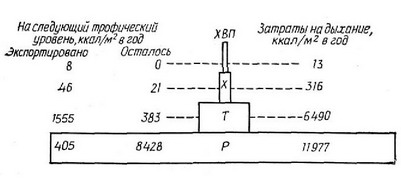
Рис. 8.5. Пирамида энергии для Силвер-Спрингс, Флорида. Р— растения, Г — травоядные, хищники, ZJJZT — хищники высшего порядка. (Из Phillipson, 1966 но Odum.)
Пирамида численностей и пирамида биомассы дают нам еще два типа экологических «пирамид». Они строятся на основе мгновенных измерений, а не скоростей и не содержат размерности времени (единицы пирамиды энергии — это кал/м в год, единицы пирамиды численностей — это число особей/м а единицы пирамиды биомассы — это г/м). Пирамида численности отражает плотность особей на каждом трофическом уровне, пирамида биомассы — биомассу (обычно измеренную в граммах сухого веса) на 1 м или на 1 м. Пирамиды численности и биомассы характеризуют только урожай на корню на данном трофическом уровне (количество в определенный момент), но не скорость оборота. Из-за отсутствия размерности времени эти пирамиды могут быть перевернутыми, т. е. низкие трофические уровни могут иметь меньшую плотность и (или) биомассу, чем высокие. Например, на одном дереве может жить множество насекомых (перевернутая пирамида численности); точно так же высокая скорость оборота дает возможность жертвам, имеющим низкую биомассу, снабжать пищей хищников, имеющих высокую биомассу, но низкую скорость оборота. Подобная перевернутая пирамида биомассы свойственна многим водным экосистемам, в которых первичные продуценты (фитопланктонные водоросли) невелики по размерам и быстро делятся, а потребляющие их зоопланктеры крупнее и живут дольше (рис. 8.6). 
Смотрите также
Особенности природоохранной деятельности в России.
В нашей стране на первом этапе становления хозяйственного
механизма природопользования недостатки административной системы руководства
проявлялись более явно и отчетливо, чем в других странах. ...
Морфофункциональные особенности лейкоцитов млекопитающих, разводимых в неволе
в условиях европейского севера
Представлены данные о морфофункциональных особенностях лейкоцитов крови различных
видов животных из отряда Carnivora— норок, песцов, лисиц и енотовидных собак, разводимых
в неволе в условиях Карел ...
Заключение.
Основным критерием экологической политики вообще является ее
действенность. Распространено мнение, что в
экологической сфере наиболее действенным является административный подход, то есть подход рас ...