Продуктивность сообществ и жизненные стратегии водорослейЭкология морского бентоса / Сообщества / Продуктивность сообществ и жизненные стратегии водорослейСтраница 3
Наивысшие показатели первичной продукции демонстрируют почти исключительно зелёные и красные эфемероиды, а из бурых — лишь нитчатые формы, т. е. виды-оппортунисты. Ламинариевые, формирующие гигантские «морские леса», т. е. дающие колоссальную биомассу, по продуктивности сильно уступают этой мелочи. Таким образом, соотношение продукции и биомассы (P/B-коэффициент) существенно выше у видов с небольшими тонкими примитивно устроенными талломами. Чистая продукция различается у перечисленных групп на порядки: для первой (Ulva, Petalonia, Porphyra) — 5,16 мг С/г в час (при чём максимальные значения достигают 10-11); для второй (Cla-dophora, Chaetomorpha, Ceramium) — 2,47; для третьей (Codium, Laurencia, Gigartina) — 1,3; для четвёртой (Padina, Gracilaria, Sargassum, Fucus, Laminaria) — 0,76; для пятой (Corallina, Jania, Amphiroa) — 0,45 и для шестой (Neogoniolithon, Lithothamnion, Hydrolithon, Peyssonnelia) — всего лишь 0,07 мг С/г в час, что в 74 раза меньше, чем у первой группы (Littler, Arnold, 1982). Аналогичное исследование, проведённое в тропических водах (на барьерном рифе у побережья Белиза), дало несколько отличающиеся результаты: для первой группы — 5,06 мг С/г в час; для второй — 5,65; для третьей — 1,09; для четвёртой — 0,88; для пятой — 0,18 и для шестой — 0,02 мг С/г в час (Littler et al., 1983) (рис. 3.49). Несмотря на инверсию результатов в двух первых, самых продуктивных группах, основная тенденция осталась прежней. И даже полученные значения для групп близки, несмотря на разные океаны и климатические зоны.
Почему так получается? Дело в том, чго продуктивность растений (и макроводорослей в том числе) связана с таким параметром, как удельная площадь поверхности (УПП) фотосинтезирующих органов — S/W, где S — площадь этих органов, а W — их масса. У водорослей, которые не только фотосинтезируют, но и поглощают питательные вещества поверхностью слоевища, эта зависимость еще больше, чем у высших растений. Наземные ботаники предпочитают обратный показатель W/S, называемый удельная поверхностная плотность. К.М. Хайлов (1988) на живых талломах ряда водорослей и на их гипсовых моделях показал, что параметр S/W имеет решающее влияние как на физико-химический обмен моделей, так и на углеродный обмен живых растений с окружающей их движущейся водой. При этом интенсификация обмена при постоянных условиях достигается ветвлением таллома.
Разработаны методики определения УПП, основанные на разделении слоевищ водорослей на цилиндрические и пластинчатые элементы, площадь поверхности которых определяется по известным формулам. Практически всё многообразие морфологии талломов можно свести к двум основным типам — осевому и пластинчатому, а также к разным их сочетаниям. Очевидно, что суммарная площадь поверхности сильно разветвлённых талломов выше, чем у разветвлённых слабо. Именно такими талломами обладает абсолютное большинство нитчаток — видов-оппортунистов. Пластинчатые талломы наращивают площадь своей поверхности за счёт рассечения пластины, образования на ней дочерних выростов — пролификаций, образования гофрировки, воланов, складчатости. Все эти структуры увеличивают площадь контакта таллома с водой и, соответственно, интенсивность обмена веществ, что подтверждено экспериментально в природных условиях (Хайлов и др., 1992).
Методика таких натурных экспериментов не имеет аналогов в мировой практике и универсальна во всех случаях, когда нужно измерить локальное движение воды — в зарослях водорослей или вокруг единичного растения, на каких-либо конструкциях и т.д. Чтобы учесть интенсивность движения воды вокруг конкретных точек слоевища или субстрата, используют гипсовые шарики определённой массы (10-50 мг). Гипсовую смесь наносят на тонкие проволочки или синтетические нити, в два-три приёма наращивая шарики нужного размера (рис.3.50). На слоевище бурой Desmarestia (длиной до 60 см) удавалось разместить до 100 шариков, на слоевище красного Ceramium — 8-10 штук. Естественно, все шарики должны быть помечены — цветом или номерами. Скорость растворения гипса при определённых скоростях движения воды известна, поэтому по начальной (до эксперимента) и конечной (после эксперимента) массе шариков можно судить об интегральной подвижности воды во время экспозиции. Для той же цели используют и модели талломов, покрытые гипсовым слоем (Хайлов и др., 1988). Эта методика позволила выяснить, что вода гораздо активнее движется у разветвлений и неровностей талломов, а также у выступов и перегибов субстрата, образуя турбулентные потоки и обеспечивая постоянный подток питательных веществ. Именно поэтому на неровностях субстрата растительность всегда богаче и мощнее, чем на ровном дне, а разветвлённые талломы имеют более интенсивный обмен со средой и, соответственно, более продуктивны. 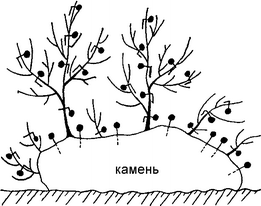
Смотрите также
Взаимодействие климата и растительности
Климат — главный фактор, определяющий характер растительности. Растения в свою
очередь также в некоторой степени воздействуют на климат. Как климат, так и растительность
оказывают решающее влияние ...
Заключение.
Основным критерием экологической политики вообще является ее
действенность. Распространено мнение, что в
экологической сфере наиболее действенным является административный подход, то есть подход рас ...
К популяционной организации политипического вида (на примере рыжей полевки -
clethrionomys glareolus shreb.)
Анализ популяционной организации и динамики численности европейской рыжей полевки
- типичного представителя мегаареальных политипических видов млекопитающих Палеарктики
- подтвердил высказанное на ...