Возникновение и эволюция рифовых сообществЭкология морского бентоса / Сообщества / Возникновение и эволюция рифовых сообществСтраница 2
Фактически третий период существования биогенных рифов начался со среднего триаса. До этого, в раннем триасе, рифовые постройки, по-видимому, отсутствовали. Сначала основную роль в образовании рифов играли водоросли и губки, реже полихеты с известковыми трубками и известковые мшанки, но в середине триаса возникли склеракти-нии, вероятно из ругоз, утративших ранее известковый скелет, из-за чего их остатки не сохранились. В триасе они вновь приобрели способность к его постройке (Келлер, Космынин, 1997). Постепенно, к позднему триасу, ведущая роль перешла к склерактиниям. В конце триаса наступает новый перерыв в рифообразовании. В этот раз он был почему-то очень краток, на порядок короче других. А восстановление рифовых сообществ произошло быстрее, чем в остальных случаях. В самом конце триаса рифы вновь получили развитие. Сначала их основными строителями были склерактинии и водоросли. В мелу на мелководье ведущая роль в строительстве каркасов рифов переходит к рудистам (рис. 3.67) — вымершей ныне группе двустворчатых моллюсков, ограниченной в своём распространении Тетисом. Рудисты обитали и на больших глубинах, но не создавали там рифов.
верхняя створка 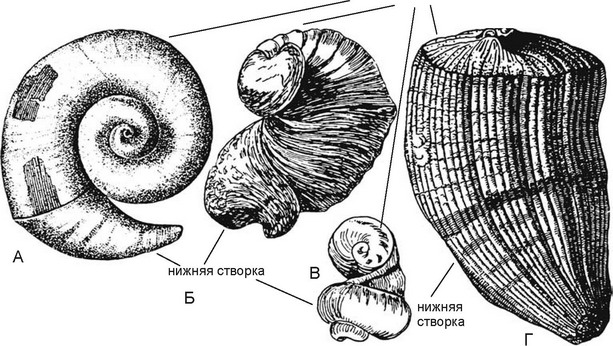
Рис. 3.67.
Рудисты.
Рудисты были одиночными ностоянно нрикренлёнными к субстрату двустворчатыми моллюсками с неравнос-творчатой раковиной, размером до 2,0 м при 0,6 м в диаметре со стенками толщиной до 10 см. Возникнув в поздней юре, в мелу они дали вспыщку таксонообразования (известно около 1000 видов), но полностью вымерли к началу кайнозоя. Позднеюрские и раннемеловые рудисты имели нреимущественно спирально закрученные створки, к субстрату прикреплялась маленькая створка или большая. Аналогом рудистов являются современные Chamidae (к прикреплённому образу жизни перешли недавно и не имеют симбионтов) и устрицы Ostreidae. По-зднемеловые рудисты были нреимущественно конические с маленькой конической створкой. Рудисты не были так чувствительны к условиям среды, как кораллы. Обитали исключительно в тропиках, где образовывали рифы, неустунавшие но масштабности рифам склерактиний.
Поскольку рудисты являются одиночными организмами, а не колониальными, как склерактинии, то образованные ими рифы имели скорее вид зарослей, но не коралловых рифов. «В течение мелового времени разнообразие рудистов возрастало за счёт развития новых адаптивных типов (пряморастущих, планоспирально завитых, лежавших свободно на субстрате, устрицеподобных и пр.); возникали скульптурированные формы, рёбра и гребни, раковины которых способствовали прочному соединению прямостоящих форм; некоторые формы цементируются к соседним особям. у многих форм верхняя створка становится прободённой или покрывается мантийной тканью, что облегчало симбиотиче-ское отношение с зооксантеллами» (Невесская, 1999: 207). Глубже склерактинии сохраняли свою ведущую роль. К концу мела рудисты вымерли и основными рифостроителями вновь стали склерактинии. Но это продолжалось недолго.
В конце мела: палеоцене,— происходит очередной перерыв в рифообразовании. На отмелях и мелководьях карбонатных платформ, где в меловое время были развиты рифовые структуры, в первой половине палеоцена они исчезли. Затем начали формироваться небольшие постройки в основном из кораллиновых водорослей и лишь местами — из губок и склерактиний (Bryan, 1991). И только в олигоцене вновь возникли сложно построенные коралловые рифы, каркасостроителями которых были кораллы, кораллиновые и зелёные водоросли, мшанки и некоторые фораминиферы, двустворки и гастроподы, но постепенно ведущая роль опять переходит к склерактиниям. Современный облик коралловые рифы приобретают в плиоцене - миоцене. Тропическая флора известковых красных водорослей (и, очевидно, растительность) существенно изменилась в начале неогена (Перестенко, 1994), т. е. одновременно с началом формирования коралловых рифов. На рис. 3.22 показан средний возраст современных родов Scleractinia. Однако, что касается родов, рифообразующих Scleractinia, то они возникли не ранее олигоцена, причём обитающие в Атлантике роды отнюдь не старше Индо-Пацифических, как это может показаться из данного рисунка. По-видимому, коралловые рифы возникли в Тетисе, затем после разделения Тетиса произошло независимое образование родов и видов в Атлантике и Индо-Па-цифике, приведшее к современной биогеографической картине.
Таким образом, рифовые сообщества формировались в течение фанерозоя неоднократно. Многие рифостроители являются фильтраторами эпифауны. Но наиболее крупные рифовые постройки возводили и возводят фотосинтетики: известковые водоросли (кембрийские строматолиты, позднее — красные и зелёные водоросли) или фотосимбио-трофные животные, впервые появившиеся, вероятно, уже в кембрии: археоциаты в кембрии, строматопороидеи (средний ордовик - пермь), табуляты (ордовик - девон), табуляты, некоторые брахиоподы в перми, фузулиниды в карбоне и перми, рудисты и ряд других двустворок (пермь-?силур-? поздняя юра - в настоящее время) и склерактинии (средний триас - ныне). Существенную роль в заполнении полостей в рифе играли также обладавшие симбионтами фораминиферы-фузулиниды (карбон - пермь), а в современных рифах — известковые водоросли. Т. е. основой функционирования мелководных рифовых сообществ всегда был фотосинтез.
Смотрите также
Введение
Исследований, непосредственно посвященных популяционной структуре вида и закономерностям
динамики численности рыжей полевки, немного, к тому же все они носят региональный
характер и основываются н ...
Биогеография морского бентоса
в биосфере Земли можно выделить четыре основных типа сравнительно независимых
друг от друга ЦС: морские, пресноводные и наземные. В свою очередь морские ЦС можно
разделить на бентосные, биотоп кот ...
Экологическая ниша
Понятие ниши пронизывает все сферы экологии. Если бы термину «экологическая ниша»
не придавали так много самых разных значений, то экологию можно было бы определить
как науку о нишах. Многие аспек ...