Причины эндогенного развития ценотических системЭкология морского бентоса / Синэкология / Причины эндогенного развития ценотических системСтраница 1
Выше мы рассмотрели процесс образования новой ЦС. Её целостность во времени поддерживается постоянством набора ниш и постоянством видового состава. И то, и другое достаточно эффективно регулирует ЦС. Таким образом, ЦС сама по себе, казалось бы, не имеет внутренних причин для развития после того, как она сформировалась. С.М. Разумовский даже считал, что ЦС «или исчезает, или остаётся в неизменном виде. О плавном эволюционировании . не может быть и речи» (Разумовский, 1999: 156). Тем не менее, на протяжении истории Земли одни ЦС сменяли другие, на их смену, в свою очередь, приходили третьи . Основная причина этого, конечно, не в непрерывной бомбардировке поверхности Земли метеоритами. Однако, ни в учебниках экологии, ни в учебниках теории эволюции причины эволюции экосистем обычно вовсе не рассматривают, или ограничиваются общими словами о росте приспособленности видов.
Достаточно ясна лишь стадия формирования ЦС. ЦС формируется для более эффективного использования имеющихся ресурсов. Пока находятся неиспользуемые ресурсы, 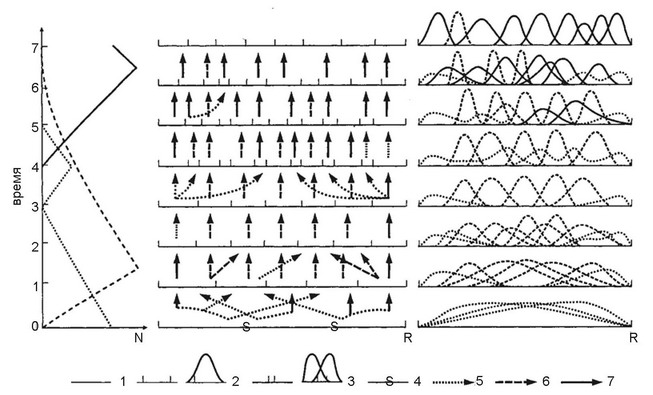
Рис. 4.39.
Схема сопряжённой эволюции сообщества и биоты, её таксономического и экологического разнообразия. Типичная носледовательность фаз филоценогенеза показана на схеме тремя разными способами в виде: динамики таксономического разнообразия (левая часть схемы); разделения лицензионного пространства и характера нреемственности (стрелки) между адантивными зонами (средняя часть схемы); плотности их упаковки (правая часть схемы). Лицензионное пространство сообщества (1); адаптивные зоны показаны двумя способами: отрезками (средняя часть схемы) или в виде распределений (правая часть схемы) в лицензионном пространстве (2); интерзональные экологические лакуны показаны промежутками между адаптивными зонами (3), нечётко выраженная граница адаптивных зон (4), слабо снециализированные таксоны показаны нунктирными линиями и стрелками (5); снециализированные таксоны 1-й формации — нрерывистыми линиями и стрелками (6), 2-й формации — сплощными линиями и стрелками (7); мера таксономического разнообразия (К); последовательность моментов (Т1-Т8) геологического времени (7). По Каландадзе, Раутиану (1993а).
происходит видообразование. Основное направление эволюции — прогрессивная специализация ассоциаций, сукцессионных рядов и т. д. Для ЦС это означает рост числа ниш (видов, ассоциаций, сукцессионных рядов и т. п.), плотности их упаковки и уровня коа-даптации. Соответственно, при формировании ЦС уменьшается объём ниш видов и сообществ и степень их перекрывания. Однако, данный процесс не может продолжаться бесконечно: ни объём используемого ресурса, ни численность популяции, ни площадь, занимаемая ассоциациями, не могут стать сколь угодно малыми. Это объясняет причину резкого замедления темпов эволюции, но не помогает понять причину последующих изменений ЦС.
H.H. Каландадзе и А.С. Раутиан (1993а,б), анализируя юрский экологический кризис блока наземных тетрапод, выдвинули гипотезу (рис. 4.39), согласно которой причина эндогенного развития ЦС заключается в следующем. Пока ЦС формируется, направление эволюции видов, входящих в её состав (филогенез), и самой системы (филоценогенез) совпадают. Направление филогенеза — прогрессивная специализация видов. Рост специализации приводит не только к увеличению эффективности потребления видом ресурса, но и к уменьшению потока энергии через популяцию данного вида и к уменьшению его численности, накоплению в системе неиспользуемых ресурсов, а также к росту чувствительности к колебаниям условий его жизни. Поскольку вероятность вымирания любого вида отлична от нуля, рост специализации приводит к увеличению этой вероятности. По достижении некоторого порога степени дробления для сообщества становится невыгодным дальнейшая специализация его членов. Вид же в силу особенностей процесса эволюции стремится к дальнейшей специализации и не может деспециализироваться. В результате дальнейшей специализации видов появляются неиспользуемые ресурсы, которые авторы теории называли «межнишевые лакуны».
Смотрите также
Основные функции и принципы экологической политики.
Комплексный характер экологических проблем
требует комплексного государственного управления в области охраны окружающей
среды. Ниже перечислим функции такого управления.
* Экологическое
прогно ...
Роль изоферментов лактатдегидрогеназы в адаптациях млекопитающих Карелии
В экстрактах тканей сердца, почек, скелетных мышц, печени, легких, селезенки
у американской норки (Mustela vison L.), песца (Aiopex iagopus), лисицы ( Vuipes
vuipes L.), лесной куницы (Martes mart ...
Особенности природоохранной деятельности в России.
В нашей стране на первом этапе становления хозяйственного
механизма природопользования недостатки административной системы руководства
проявлялись более явно и отчетливо, чем в других странах. ...