Причины эндогенного развития ценотических системЭкология морского бентоса / Синэкология / Причины эндогенного развития ценотических системСтраница 2
Отдельных ресурсов недостаточно для поддержания популяции специалиста, но суммарно разные ресурсы достаточны для поддержания популяции генералиста (неспециализированного вида). Источником генералис-тов выступают экотоны. Появление в сообществе таких генералистов представляет определённую опасность для сохранения прежней структуры системы. Если структурирующие возможности системы достаточно велики, то на базе этих генералистов возникает новая формация специалистов, в общих чертах воспроизводящая организацию прежней, и устойчивость системы восстанавливается. По мере роста числа специалистов второй формации насыщенность сообщества нишами увеличивается, со временем вновь переходит оптимальный уровень, и цикл повторяется. Модель описывает сообщество, состоящее исключительно из специалистов, причём уровень их специализации примерно одинаков и постоянно растёт. Такое сообщество, если и возможно, отнюдь не является типичным. В сообществах в норме сосуществуют специалисты и генералисты, а уровень специализации видов различен и не может расти бесконечно. 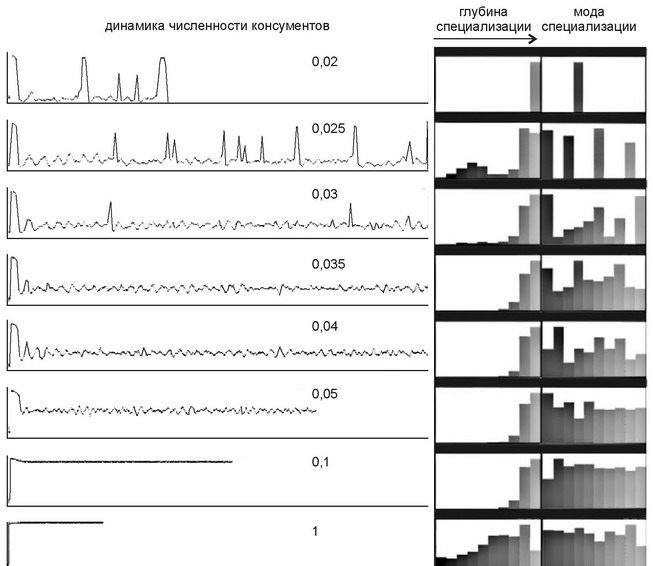
Рис. 4.40.
Динамика численности консументов в зависимости от продуктивности. Цифры — поток энергии через сообщество. Справа: типичные «портреты сообществ». Глубина специализации имеет 10 градаций; мода снециализации отражает нотребление 10 различных ресурсов. По Грабовскому (2005).
Эту же проблему изучал В.И. Грабовский (2005) на модели, построенной на основе идеологии клеточных автоматов (рис. 4.40). В отличие от объяснения post factum, на модели легче выделить необходимые и достаточные причины, поскольку в модели условия можно контролировать, а эксперименты — повторять. Такая модель должна:
1. Включать минимум допущений (правило «бритвы Оккама»*), которые не противоречили бы имеющимся данным (как палеонтологическим, так и неонтологическим).
2. Демонстрировать поведение системы, которое находится в фокусе наших объяснений: длительные периоды когерентной эволюции, сменяющиеся короткими периодами перестроек (некогерентной эволюции).
В модели рассмотрен простейший случай сообщества, состоящего из одного трофического уровня, виды которого могут специализироваться, и десяти видов ресурсов, пополняемых с постоянной скоростью, т. е. моделировалось сообщество, аналогичное рассмотренному H.H. Каландадзе и А.С. Раутианом (1992, 1993). Изучение модели показало, что с увеличением потока энергии через сообщество (роста продуктивности и/или площади, занятой сообществом), стабильность сообщества растёт. При снижении потока энергии в системе постепенно накапливаются неиспользованные ресурсы. В модели H.H. Каландадзе и А.С. Раутиана неиспользуемые ресурсы появлялись в результате сужения ниш видов из-за прогрессивно увеличивающейся их специализации. Согласно их модели, специализация является как бы самоцелью (направление филогенеза — прогрессивная специализация видов). В результате роста специализации входящих в сообщество видов, эффективность использования всем сообществом имеющихся ресурсов снижается, что эквивалентно утверждению о снижении эффективности (полноты) использования имеющихся ресурсов (т. е., в конечном счете, приспособленности) каждым из видов сообщества. Существование такого процесса специализации, который имел бы результатом снижение эффективности использования видом имеющихся ресурсов представляется весьма сомнительным. В модели же В.И. Грабовского накопление ресурсов происходит в результате недостаточно эффективной специализации. Специализация при этом, в противоположность модели H.H. Каландадзе и А.С. Раутиана (1992, 1993) растёт не безгранично, а только до тех пор, пока растёт эффективность использования ресурсов. Базовые условия модели В.И. Грабовского гораздо больше походят на правду, поскольку специализация, с одной стороны, не может расти неограниченно (как предполагают H.H. Каландадзе и А.С. Раутиан), а с другой — всегда недостаточно эффективна. Это происходит из-за того, что (1) непрерывно меняющиеся условия изменяют вектор эволюции и (2) ни одна из ЦС не состоит на 100% из эндемов, какая-то часть видов, зачастую существенная, входит в состав других ЦС. Направление специализации в разных ЦС неизбежно различается, что снижает эффективность использования ресурсов в каждой из ЦС, в которую входит такой вид.
Смотрите также
Экономические методы охраны окружающей среды и особенности их использования в России
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Введение
Исследований, непосредственно посвященных популяционной структуре вида и закономерностям
динамики численности рыжей полевки, немного, к тому же все они носят региональный
характер и основываются н ...
Роль изоферментов лактатдегидрогеназы в адаптациях млекопитающих Карелии
В экстрактах тканей сердца, почек, скелетных мышц, печени, легких, селезенки
у американской норки (Mustela vison L.), песца (Aiopex iagopus), лисицы ( Vuipes
vuipes L.), лесной куницы (Martes mart ...