Методы группирования регионовЭкология морского бентоса / Биогеография — общие вопросы / Методы группирования регионовСтраница 3
«Хотя понятия «саванна» и «степь» были введены для обозначения определённых типов растительности, в последнее время они применяются географами скорее для обозначения соответствующих климатических зон_ В этом случае, по мнению автора, следовало бы пользоваться чисто климатической терминологией ботанические термины должны применяться для характеристики какой-то определённой категории растительного покрова» (Вальтер, 1968: 276). 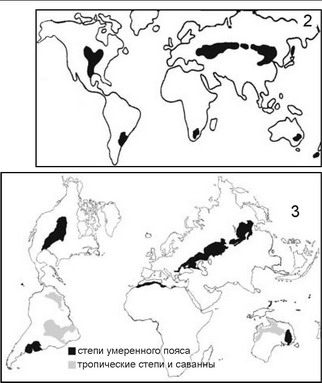
выделения регионов и в географии весьма субъективны. Это следует хотя бы из того, что один и тот же биом разные авторы понимают весьма различно, хотя и называют одинаково. В качестве примера на рис. 5.28 дано распространение биома степей по разным авторам. Из сравнения данных схем становится очевидной высокая степень их субъективности, неоднозначности применения этого подхода в самой географии.
Та же картина наблюдается и в любом более мелком масштабе, в том числе и в масштабе ландшафтов. Например, болото — вполне ясный и хорошо очерченный, казалось бы, тип ландшафта. Однако, даже по официальным данным разных ведомств площадь болот в России различается на 20%: в начале 1998 г. в лесном фонде было 128,3 млн. га (Лесной фонд России, 1999), но по данным государственного учёта земель болота в лесном фонде России занимают всего 101,2 млн. га (Думнов и др., 2001). Неофициальные данные дают ещё больший разброс: до 273,0 млн. га (Ефремов и др., 1994) и даже 361,1 млн. га (Вомперский и др., 1999). Достойно удивления, что при общем разбросе размеров площадей (в три раза) все источники считают возможным приводить свои цифры с точностью до четырёх значащих цифр.
Из сказанного выше вовсе не следует, что зональная биогеография не нужна и бессмысленна. Однако, для получения осмысленных с биологической точки зрения зон объединять следует биогеографические регионы, выделенные на основе тех свойств, которые изучают. В биологических исследованиях это означает, что следует использовать биологические критерии тем более, что имеются образцы такого районирования с получением биологически значимой информации.
В качестве примера можно привести выделение зон наземной растительности С.М. Разумовским (1999). Начав с установления ботанико-географических провинций, он выявил коренные ассоциации их климаксов, затем сравнил облик древесных эдификаторов климаксных ассоциаций, общее число которых (для всей Земли) составляет лишь несколько десятков, и обнаружил, что «эти виды очень легко и естественно распадаются на четыре различных между собой и очень монолитных морфологических типа:
1. Вечнозеленые перистолистные деревья с почками, лишёнными почечных чешуй.
2. Вечнозеленые широколистные деревья с почками, защищёнными почечными че-шуями.
3. Летнезелёные широколистные деревья с почечными чешуями.
4. Вечнозелёные деревья с игольчатыми листьями ксероморфной структуры и с почечными чешуями» (Разумовский, 1999: 73). 
Рнс. 5.28.
Распространение биома степей по
разным авторам. 1 — по: А.Г. Исаченко, А.А. Шляпникову (1989); 2 — по: В.Г. Мордковичу (1982); 3 — по классификации ЮНЕСКО (План действий ., 1984).
Эти зоны и являются естественными биологическими зонами растительности Земли: тропическими, субтропическими и широколиственными лесами и тайгой.
ЦС, входящие в одну зону, могут приобрести своё сходство чисто конвергентно и вообще не иметь ничего общего ни в своём составе, ни в происхождении. Сходная ситуация имеется и в таксономии: в воде активно плавающие животные приобретают форму рыбы, но гораздо информативнее классификация не на основе жизненных форм, а филогенетическая, учитывающая то, что одни из этих «рыб» — рыбы, другие — рептилии, третьи — млекопитающие, четвёртые — птицы, пятые — головоногие моллюски и т. д. Из того факта, что в воде они приобрели сходную форму, мало что можно сказать об их внутреннем строении, биохимии, происхождении и т. п. Но все эти данные вполне доступны из филогенетической классификации, построенной на основе их генетического родства. Аналогично, объединение биогеографических регионов по их происхождению гораздо более информативно для понимания их биологических свойств, тогда как зональные классификации лишь показывают многообразие ЦС, образовавшихся на разной биотической основе, но в сходных абиотических условиях.
Для других целей (например, для оптимального размещения санаториев) более информативными являются иные логические основания для объединения регионов, разные для различных задач. В некоторых случаях более информативными будут и чисто физиономические основания.
Смотрите также
Экологическая ниша
Понятие ниши пронизывает все сферы экологии. Если бы термину «экологическая ниша»
не придавали так много самых разных значений, то экологию можно было бы определить
как науку о нишах. Многие аспек ...
Экономические механизмы охраны природы
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Платежные инструменты
Платежные инструменты - это правовые средства осуществления переводов денежных средств (наличных и безналичных) с целью совершения платежа.
Порядок выполнения операций с применением платежных инст ...