Структура и динамика популяцийКниги по екологии / Экология / Структура и динамика популяцийСтраница 10
Итак, при анализе природных совокупностей особей (популяций) необходимо различать понятия: частота гена (количественное соотношение аллелей одного какого-либо локуса), частота генотипа (количественное соотношение разных генотипов) и частота фенотипа (количественное соотношение разных фенотипов).
Формула Харди—Вайнберга пригодна лишь для предельно упрощенной ситуации, для идеальной бесконечно большой популяции (иногда ее называют «менделевской») и при отсутствии давления каких-либо факторов. Кроме того, как известно, на частоту фенотипов оказывает влияние не только частота данного аллеля, но и такие его свойства, как доминантность, рецессивность, пенетрантность и экспрессивность. Таким образом, при анализе природных популяций данная формула применима лишь с большими оговорками.
Существование двух (или более) генетически различных форм в популяции в состоянии длительного равновесия в таких соотношениях, что частоту даже наиболее редкой формы нельзя объяснить только повторными мутациями, называется полиморфизмом. В качестве примера полиморфизма можно привести три формы цветков у примулы (Primula vulgaris), рис. 9.8.
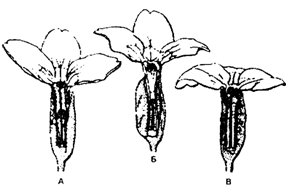
Рис. 9.8. Три формы цветков у примулы (Primula vulgaris),
из Ф. Шеппарда, 1970
В природных популяциях примулы (Primula vulgaris) всегда есть особи с длинным пестиком и короткими пыльниками (А), длинными пыльниками и коротким пестиком (Б) и одинаковыми по длине пыльниками и пестиком (В). Самоопыление возможно только у цветов типа В. Гетеростилия способствует перекрестному оплодотворению.
Полиморфизм по механизму возникновения и поддержания разделяется на две большие группы: гетерозиготный и адаптационный.
Гетерозиготный полиморфизм устанавливается в результате давления на популяцию естественного отбора, положительно отбирающего гетерозигот.
Адаптационный полиморфизм — это две или несколько генетически различных форм внутри популяции, подвергающихся положительному отбору в разных экологических условиях.
В качестве примера можно привести адаптационный полиморфизм в популяции двухточечной тлевой (божьей) коровки Adalia bipunctata (рис. 9.9).
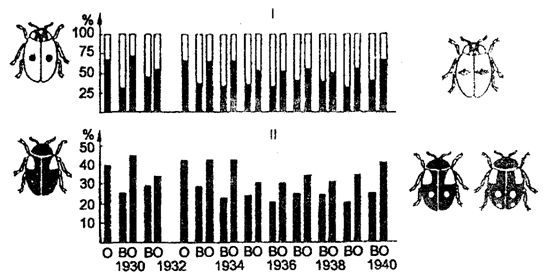
Рис. 9.9. Адаптивный полиморфизм в популяции тлевых
(божьих) коровок Adalia bipunctata:
I — процентное содержание черной и красной форм при весеннем (В) и осеннем (О) сборах; II — частота доминантного гена А (черная окраска) в популяциях весной и осенью каждого года (в%) (по Н. В. Тимофееву-Ресовскому и Ю. М. Свирежеву, 1965)
На протяжении 10 лет в изучении популяции тлевой коровки осенью — при уходе на зимовку — черных форм было от 50 до 70%, а весной — при выходе из зимовки — от 30 до 45%. Красных форм осенью было меньше 50%, а весной — больше. Красные формы, как было установлено, лучше переносят в зимний период холод, а черные — интенсивнее размножаются летом. Отсюда отбор направлен на сохранение большего числа красных жуков зимой и черных — летом. Разнонаправленное давление отбора в отдельные периоды жизни популяции способствует выработке устойчивого адаптационного полиморфизма.
Положение о генетическом единстве популяции является одним из наиболее важных выводов популяционной генетики: любая популяция представляет сложную генетическую систему, находящуюся в динамическом равновесии.
9.8. Рост популяций и кривые роста
Если рождаемость в популяции превышает смертность, то популяция, как правило, будет расти. С увеличением плотности скорость роста популяции постепенно снижается до нуля. При нулевом росте популяция стабильна, т. е. размеры ее не меняются. Отдельные организмы при этом могут расти и размножаться. Нулевая скорость роста означает лишь то, что скорость размножения, если оно происходит, уравновешена смертностью. Данная картина характерна для ряда одноклеточных и многоклеточных организмов, например для клеток водорослей в культураль-ной жидкости, для фитопланктона озер и океанов весной, для насекомых (мучные хрущаки, а также клещи, интродуцирован-ные в новое местообитание с обильными запасами пищи, где нет хищников).
Смотрите также
Биологические опасности, связанные с пищей
Научно-технический прогресс сильно повлиял на сферу производства продуктов питания. Технологическая обработка продуктов, консервирование, рафинирование, длительное и неправильное хранение резко снизил ...
Экономические методы охраны окружающей среды и особенности их использования в России
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...
Основные функции и принципы экологической политики.
Комплексный характер экологических проблем
требует комплексного государственного управления в области охраны окружающей
среды. Ниже перечислим функции такого управления.
* Экологическое
прогно ...