Пол, соотношение полов, половой отбор и типы брачных отношенийЭволюционная экология / Принципы популяционной экологии / Пол, соотношение полов, половой отбор и типы брачных отношенийСтраница 7
Если в популяпии равное соотношение полов и моногамная система брачных отношений, то любой индивидуум (как самеп, так и самка) имеет достаточно высокую вероятность спариться и передать следующему поколению свои гены. Однако при равном соотношении полов и полигинии избранная группа наиболее при-способленых сампов обеспечивает непропорпионально большое число спариваний. Покрытые боевыми шрамами сампы морских львов, «отбившие» скалистые островки, на которых обычно происходит спаривание, часто имеют гаремы по 10—20 самок. В данном случае немногие доминирующие сампы являются отпами подавляющего большинства потомства, и их гены составляют половину генофонда последующего поколения. Поскольку признаки, обеспечивающие их высокие боевые качества и делающие их доминантными, наследуются и переходят сыновьям, борьба за брачную территорию в следующем поколении может усиливаться. Так как лишь те сампы, которые побеждают соперников, способны передать свои гены потомству, в данном случае строго соблюдается принпип «все или ничего». В результате такой интенсивной конкуренпии за брачную территорию внутриполовой отбор приводит к ярко выраженному половому диморфизму в размерах: если вес взрослой самки обычно менее 500 кг, то вес взрослых сампов может достигать тонны. Различие в размерах полов особенно велико у калифорнийского морского льва Zalophus, самки которого весят только около 100 кг, тогда как сампы — почти 500 кг. Вероятно, предел увеличения разницы в размерах определяется всевозможными другими экологическими факторами, например такими, как давление хищников, эффективность добывания пищи и ее доступность (см. также гл. 4 и 7).
В чем-то сходная ситуапия наблюдается среди различных по-лигинныхиполибрахигинных видов птип- Полибрахигиния встречается у многих тетеревиных, сампы которых проявляют свою половую активность в группах, собирающихся на токовищах. Доминируют обычно старшие самцы, которые занимают центральную часть общего токовища и участвуют в непропорционально большой доле спаривания. Самки, находящиеся на токовище, быстра бросают периферийных самцов, стремясь к спариваниям с самцами,, занимающими центральное положение. Для многих тетеревиных характерен заметный половой диморфизм в размерах, оперении,, окраске и поведении. В дополнение к внутриполовому отбору эпи-гамный отбор, действуя через выбор, осуществляемый самками,, также может привести к возникновению и усилению полового диморфизма. Как правило, оба типа полового отбора происходят одновременно и различить их результаты часто бывает трудно.
Рис. 5.36. График, показывающий, что самцы (I) красноглазого виреона (Vireo olivaceous) предпочитают кормиться на большей высоте, чем самки (II). (Из Williamson Р., 1971. Copyright by the Ecological Society of America.) 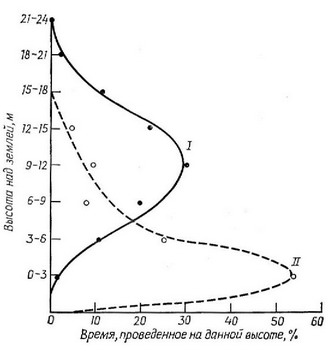
Выбор самками для спаривания ярких, бросающихся в глаза самцов, по-видимому, приводит к возникновению у самцов в процессе эволюции ряда странных признаков, таких, например, как очень длинные хвосты, характерные некоторым райским птицам. Беседковым птицам (шалашникам) удалось избежать яркости (а следовательно, и опасной заметности) путем выработки в эволюции уникальных особенностей поведения: самцы для привлечения самок и ухаживания за ними строят богато украшенные беседки, служащие как бы показателем сексуальной привлекательности самца. Весьма часто одни и те же половые признаки (такие, как размер, окраска, оперение, особенности пения и поведения) служат одновременно для демонстрации как своего положения среди представителей того же пола, так и своей привлекательности для особей противоположного пола. Подобное совпадение явно имеет эволюционный смысл, поскольку суммарная приспособленность особи определяется ее успехом, достигаемым совместным действием обоих типов полового отбора, которые между собой должны обычно положительно коррелировать. Кроме того, такое совпадение половых сигналов позволяет также экономить расход энергии.
Половой диморфизм иногда выполняет еще и другую экологическую функцию, уменьшая перекрывание ниш и снижая интенсивность конкуренции между представителями разных полов (рис. 5.36). У некоторых яшериц, обитающих на островах (Schoener, 1967, 1968а), и некоторых птиц (Selander, 1966) сильный половой диморфизм в размере ротового аппарата (челюстей и клювов) коррелирует с дифференцированным использованием п
Смотрите также
Биогеография морского бентоса
в биосфере Земли можно выделить четыре основных типа сравнительно независимых
друг от друга ЦС: морские, пресноводные и наземные. В свою очередь морские ЦС можно
разделить на бентосные, биотоп кот ...
Взаимодействие климата и растительности
Климат — главный фактор, определяющий характер растительности. Растения в свою
очередь также в некоторой степени воздействуют на климат. Как климат, так и растительность
оказывают решающее влияние ...
Экономические методы охраны окружающей среды и особенности их использования в России
Проблема защиты экологии встала перед
человечеством сравнительно недавно. Но уже в нашем веке, который ознаменовал
себя масштабным истощением природных ресурсов, огромным количеством вредны ...