Наблюдения в природеЭволюционная экология / Взаимодействия между популяциями / Наблюдения в природеСтраница 4
Подобно Бермудским островам в океане, вершины гор представляют собой «острова» наземного ландшафта и на них обнаруживают сходные явления. Например, два вида саламандр, принадлежащие к одному роду (Plethodon jordani и Р. glutinosus), — симпатрические обитатели гор восточной части С1ПА. В зоне симпатрии они разделены по высоте, при этом Р. glutinosus встречается ниже, чем р. jordani. Вертикальное перекрывание между этими видами никогда не превышает 70 метров (Hairston, 1951). На вершинах гор, где встречается Р. jordani, Р. glutinosus ограничен более низкими участками, вместе с тем на соседних горах, где нет Р. jordani, Р. glutinosus поднимается выше, нередко до самого пика.
Расширение ниши в условиях ослабления межвидовой конкуренпии было названо «экологическим высвобождением». Дальнейшие данные, свидетельствующие о конкуренпии, вытекают из логического следствия экологического высвобождения: когда материковые формы заселяют острова, местные виды нередко исчезают, по-видимому, в результате конкурентного исключения. Например, многие птипы, которые раньше встречались только на
Гавайях (эндемичные гавайские виды), исчезли вскоре после интродукции таких материковых птиц, как домовый воробей и скворец. Подобные потери понесла и австралийская фауна сумчатых (например, сумчатый волк) после вселения видов плацентарных млекопитающих (например, собаки динго и европейской лисицы). История ископаемых организмов изобилует примерами естественных нашествий и последующих вымираний. Наиболее простым и правдоподобным объяснением многих таких событий может быть конкуренция: выжившие виды имели конкурентное преимущество, а перекрывание ниш было слишком большим для сосуществования. Поэтому до того, как естественный отбор привел к смещению признаков и разделению ниш, один из видов вымирал. Множество других примеров экологических нашествий как у растений, так и у животных обсуждается в работе Элтона (Elton, 1958).
Наконец, для оценки того, играет ли конкуренция важную роль в природе, используют данные по таксономическому составу сообщества. Поскольку конкуренция у близкородственных видов должна быть особенно сильной, можно предположить, что в пределах любого данного естественного сообщества будет найдено меньшее число пар видов одного рода по сравнению с числом встречающихся в более широкой географической области различных групп видов и родов в совершенно случайной выборке. Такое небольшое число симпатрических видов одного рода, если оно обнаружилось, позволяет предположить, что конкурентное исключение более часто наблюдается среди близкородственных видов, чем среди видов, отдаленных в систематическом отношении. Это положение было проверено на многих сообществах Элтоном (Elton, 1946), который хорошо осознавал трудности, связанные с проблемой определения «сообщества». Нередко пару родственных видов могут составлять виды из разных, но граничащих друг с другом сообществ. Такие пары необходимо везде, где только можно, исключать из рассмотрения. Несмотря на подобную возможность завышения доли пар видов одного рода, Элтон обнаружил меньше видов, чем ожидалось в условиях строгого соблюдения случайности. Позднее Уильяме (Williams, 1964) показал, что анализ Элтона ошибочен, и применил правильный статистический подход к решению этой проблемы. Используя правильную методику Уильямса, Терборг и Веске (Terborgh, Weske, 1969) вычислили ожидаемое число пар родственных видов перуанских птиц в семи местообитаниях (рис. 6.15). Эти авторы обнаружили, что четыре местообитания, наиболее богатые по общему числу видов, содержали родственных видов больше ожидаемого; этот факт опроверг предположение о широком распространении конкурентного исключения у видов одного рода в рассматриваемой орнитофауне. Мне также не удалось обнаружить соответствующего обеднения пар видов, принадлежащих к одному роду, в ряде сообществ ящериц (Pianka, 1973). Было бы полезно провести более подробный анализ такого типа, использовав при этом широкий набор видов, обитающих в различных сообществах. 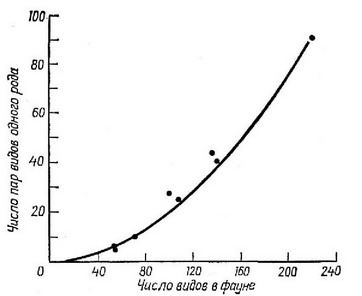
Рис. 6.15. Наблюдаемое (точки) и расчетное (кривая) число пар родственных видов птиц в 7 местообитаниях в Неру. Вся орнитофауна 7 областей состояла из 221 вида и была предс
Смотрите также
Влияние низкотемпературного закаливания на протеолитическую активность и содержание
фотосинтетических пигментов в листьях проростков озимой пшеницы
На проростках озимой пшеницы (Тп11сит aestivum 1.) изучена динамика активности
амидаз, цистеиновых про-теиназ, а также содержания фотосинтетических пигментов при
холодовом (4°С) закаливании. Показ ...
Заключение.
Основным критерием экологической политики вообще является ее
действенность. Распространено мнение, что в
экологической сфере наиболее действенным является административный подход, то есть подход рас ...
Морфофункциональные особенности лейкоцитов млекопитающих, разводимых в неволе
в условиях европейского севера
Представлены данные о морфофункциональных особенностях лейкоцитов крови различных
видов животных из отряда Carnivora— норок, песцов, лисиц и енотовидных собак, разводимых
в неволе в условиях Карел ...