Континуализм и структурализмЭкология морского бентоса / Синэкология / Континуализм и структурализмСтраница 7
В приведённых выше моделях события развиваются во времени. Предложенные для их объяснения теории также растолковывают развитие процесса во времени, хотя реальные события движутся не только во времени, но и в пространстве. Экспериментальные сосуды Г.Ф. Гаузе с точки зрения инфузорий (т. е. учитывая их способность к перемещению в пространстве) равноценны одной точке.
Между тем, существует множество работ, в которых показано, что учёт пространства может существенно изменить результаты. Проиллюстрируем это наглядной моделью, основанной на идеологии клеточных автоматов* (Грабовский, 2005). В ней особи двух или нескольких видов (число видов можно задать от двух до десяти) взаимодействуют на совершенно однородном моделируемом поле. Целиком особи всех видов имеют равную конкурентоспособность и занимают одну нишу (имеют идентичные потребности). с помощью модели можно изучить динамику во времени: (1) суммарной по полю относительной численности видов и (2) распределение отдельных видов в пространстве. Пространство измеряют в размерах особей, время — в генерациях. Измерения этих параметров в относительных, а не абсолютных величинах (метры, сутки и т. п.) биологически более правильно (Азовский, 2003). При размножении каждая особь делится на несколько эквивалентных ей потомков (можно задать разное число потомков). Каждый потомок расселяется в пределах некоторого задаваемого радиуса. Если выбранный участок оказывается свободным — занимает его. Если участок занят другим видом, то расселяющаяся особь с вероятностью 0,5 (виды имеют равную конкурентоспособность!) либо вытесняет его, либо вымирает. При половом размножении добавляется ещё один задаваемый параметр: радиус поиска полового партнёра (разноса гамет, пыльцы, спермиев, неоплодотворённых яйцеклеток и т. п.). Модель показывает, что виды с одинаковой конкурентоспособностью, занимающие одну нишу в однородном пространстве, могут сосуществовать весьма долго, если это пространство достаточно велико. Изменение параметров модели (радиуса расселения, радиуса поиска полового партнёра, размера поля, числа потомков) влияет на результат. На модели получено, что продолжительность сосуществования видов уменьшается при: 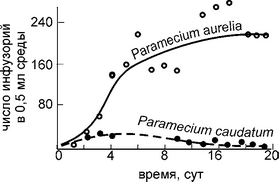
Рис. 4.3.
Динамика нонуляций инфузорий Paramecium aurelia и P. caudatum в совместной культуре По G.F. Gause, 1934 из: Ги-лярова (1990).
— уменьшении отношения размера экспериментального поля к радиусу перемещения особей (т. е. по мере приближения поля к точке), предельный вариант (поле равно точке) с максимальной неустойчивостью сосуществования видов и рассматривает теория Га-узе-Вольтерра;
— увеличении плодовитости.
При низкой способности к расселению и большом радиусе поиска полового партнёра (ситуация типичная для, например, деревьев тропических лесов, имеющих крупные плоды, которые разносятся на расстояние гораздо меньшее, чем пыльца), немалое число видов (в данной модели максимальное число — 10) с идентичными нишами способны существовать весьма долго. Прогон модели с параметрами фитопланктона (радиус расселения мал, размножение бесполое) даёт ещё более устойчивую ситуацию сосуществования 10 видов с полностью идентичными экологическими нишами. Виды образуют пятна, форма пятен довольно изменчива, положение пятен более постоянно. Получаемые на моделируемом поле пятна разных видов весьма напоминают наблюдения в природе. Такая картина получается и при изучении распределения сообществ (примерно соответствующему распределению наиболее массовых видов) инфузорий на литорали Белого моря (Бурковский, 1992, 2006: 154) (рис. 4.5). По результатам съёмок с интервалом в 2 недели (примерно 30 поколений) форма пятен сильно изменилась, но многие пятна остались примерно в тех же местах. Таким образом, учёт пространства существенно изменяет результаты. Более того, клеточные автоматы позволяют смоделировать ситуации, для которых не удаётся подобрать систему уравнений и коэффициенты. При этом получаемые результаты вполне соответствуют наблюдениям в природе не только по моделируемым свойствам, но даже и по тем, какие вовсе никак не учитывали при построении модели. Кроме уже разобранных свойств однови-довых пятен, вполне соответствует наблюдениям в природе и распределение относительных частот видов по результатам модели (рис. 4.4а). Таким образом, оказывается, что противоречие между фактическими данными и обычно используемыми математическими моделями, в частности системой дифференциальных уравнений Вольтерра,
Смотрите также
Основные функции и принципы экологической политики.
Комплексный характер экологических проблем
требует комплексного государственного управления в области охраны окружающей
среды. Ниже перечислим функции такого управления.
* Экологическое
прогно ...
Влияние низкотемпературного закаливания на протеолитическую активность и содержание
фотосинтетических пигментов в листьях проростков озимой пшеницы
На проростках озимой пшеницы (Тп11сит aestivum 1.) изучена динамика активности
амидаз, цистеиновых про-теиназ, а также содержания фотосинтетических пигментов при
холодовом (4°С) закаливании. Показ ...
Введение
Исследований, непосредственно посвященных популяционной структуре вида и закономерностям
динамики численности рыжей полевки, немного, к тому же все они носят региональный
характер и основываются н ...